Neural-network fitting as a structure sensor
Can the fitting process of a plain autoencoder act as a sensor for hidden structure in single-cell data — recovering a biological axis it was never shown?
An undercomplete autoencoder is trained only to reconstruct gene expression — it never sees donor age. If age nonetheless surfaces in its compressed code, the fit itself would be a usable sensor. Calibrated on the age axis as ground truth, the method fails its own pre-committed bar — and the way it fails is informative.
The design
One autoencoder architecture, three matched data conditions, and a pre-committed pass/fail rule fixed before any result was seen.
- Ground truth. Donor age is a donor-level variable, so every probe uses leave-donors-out cross-validation (effective n = 167 donors, not cells). A cell-level split would measure donor leakage, not an age law.
- Three conditions.
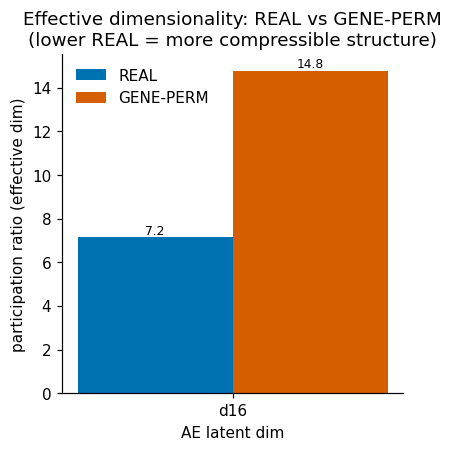
REAL(untouched) ·GENE-PERMUTED(co-expression destroyed, marginals kept — an artifact floor) · age-shuffled (a within-source permutation null for the probe). - Verdict A∧B∧C∧D, locked in advance: A age beats the within-source null · B the effect does not reproduce on gene-permuted data · C the latent beats a matched linear PCA · D the signal survives a data-source (batch) control.
- Independent cross-check against the perceptome 9-PC eigenspace — a biology-curated coordinate system built from 154 cell types.
What the fit reveals
The sensor signal is not the reconstruction loss. It is read three ways, each against a matched null.
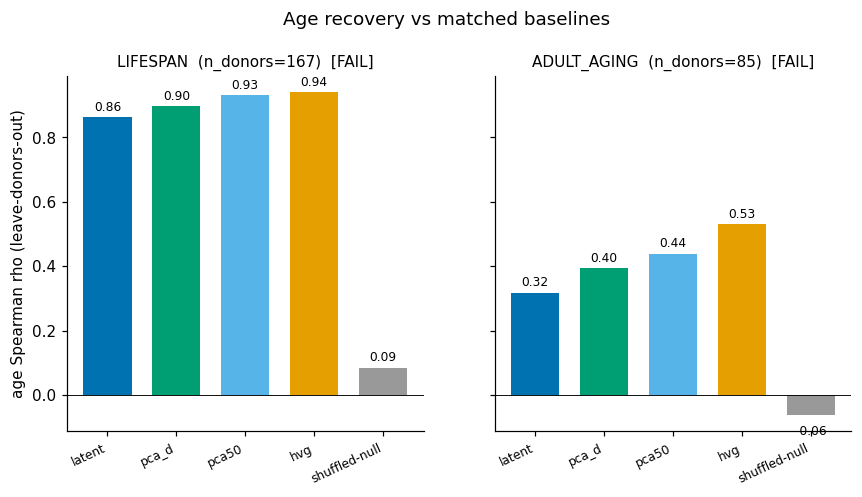
The instrument is sound — but narrowly
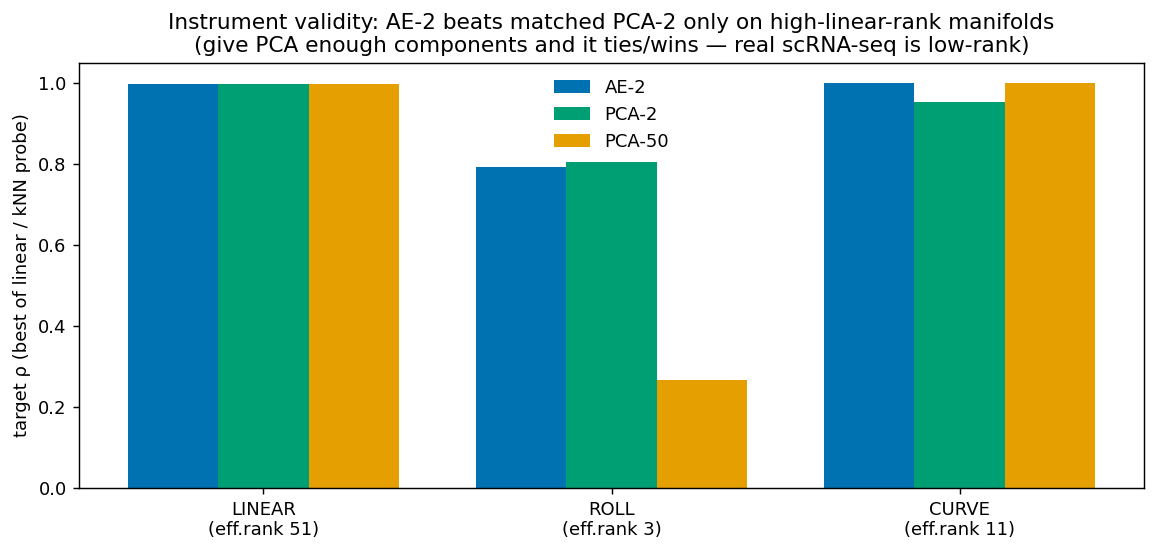
Could a linear probe be hiding a nonlinear success? A synthetic positive control settles it.
The independent cross-check
What was found
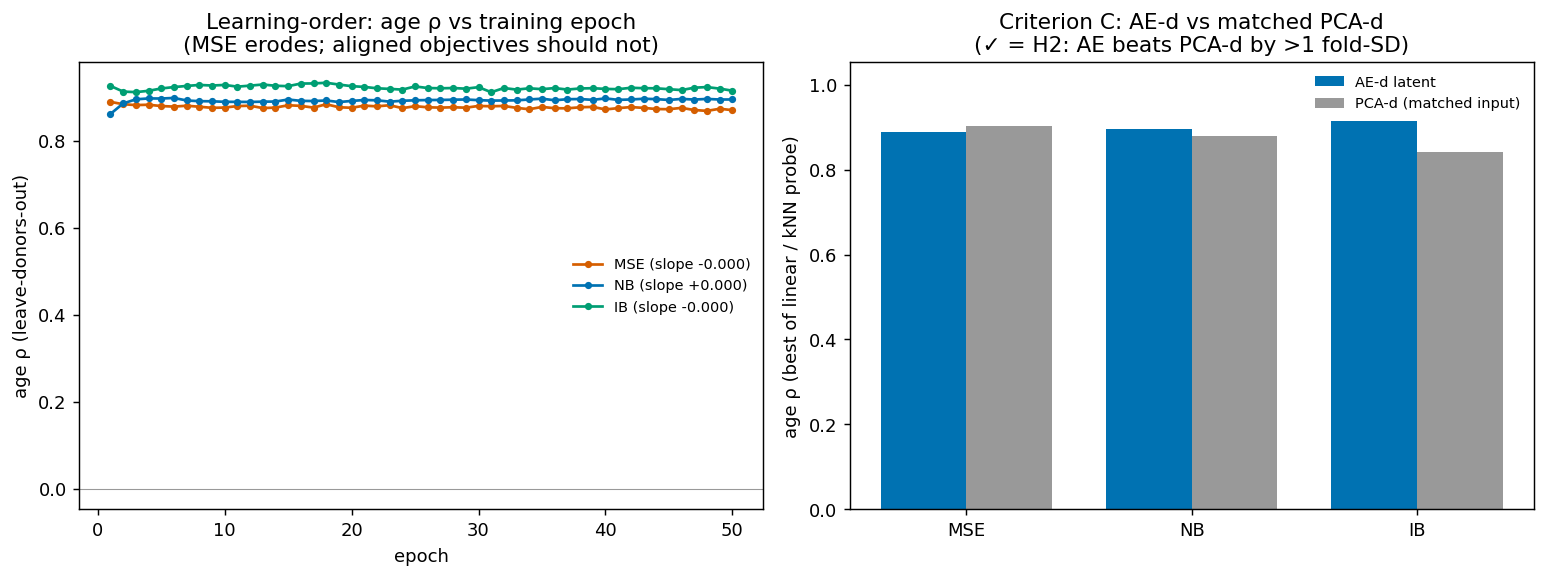
- (Negative.) The autoencoder does not beat matched linear PCA at recovering age — 0/2 across seeds × dims (lifespan latent 0.86–0.89 vs PCA 0.90–0.93; adult 0.35–0.40 vs 0.44–0.48). More capacity (d=32) does not rescue it, so the bottleneck is the data geometry, not the network size.
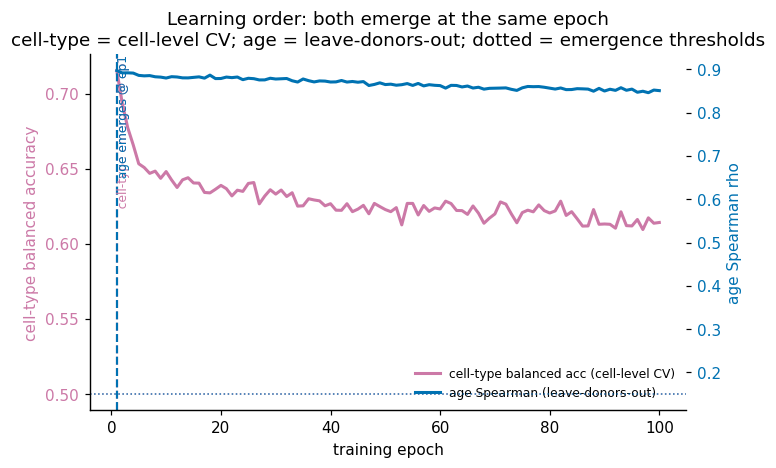
- Training erodes the signal under MSE. Age decodability is highest at initialisation (≈ random projection) and declines over 100 epochs; the unsupervised fit specialises for reconstruction and overwrites the linearly-accessible age axis.
- Naive adult-aging recovery is partly a batch artifact. The latent predicts dataset-of-origin at 0.94 balanced accuracy (chance 0.33); within a single source the adult signal collapses to ρ ≈ 0.14 (n.s.). The pre-registered source control (D) catches exactly this.
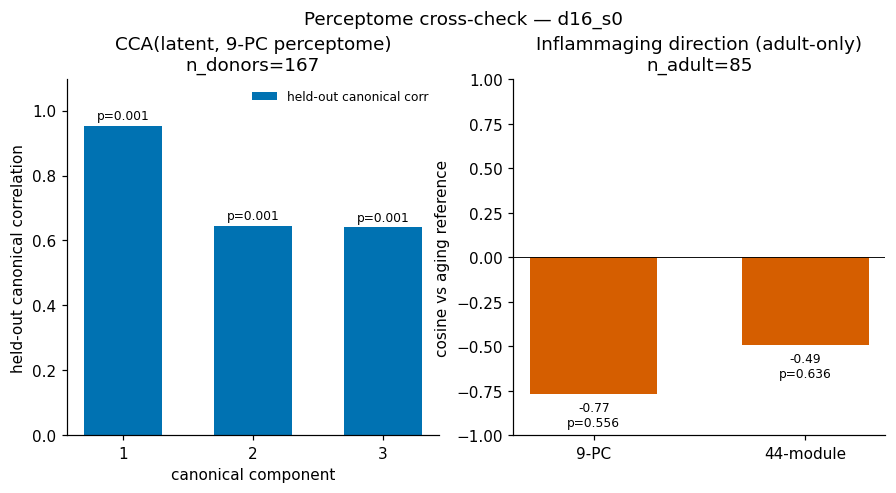
- The latent independently re-derives the perceptome eigenspace (CCA 0.95 / 0.56 / 0.74, p = 0.001) but not its aging axis (cosine to inflammaging −0.78 / −0.50, n.s.) — perceptome axes look pathology-specific.
- The instrument is sound but the regime is wrong. A positive control confirms the autoencoder can beat matched PCA — only when intrinsic-dim < bottleneck < linear-rank. Single-cell data (low effective rank, linear age) is outside that window; no objective changes that.
| Target | A · beats null | B · not on permuted | C · beats PCA | D · source-robust | Verdict |
|---|---|---|---|---|---|
| Lifespan (167 donors) | ✅ | ✅ | ❌ | ✅ | FAIL |
| Adult-aging (85 donors) | ✅ | ✅ | ❌ | ❌ | FAIL |
What this means
The result is a clean, mechanistically dissected negative. Autoencoder-fitting-as-a-sensor beats linear baselines only on data whose structure is nonlinearly entangled — high linear rank, low intrinsic dimension. Standard single-cell expression, after the usual preprocessing, is well-approximated by ~50 linear components and carries a linearly-dominated age axis; there is simply no nonlinear advantage for the bottleneck to find that a generous linear method does not already capture. The training objective controls whether the fit erodes the signal (it does under MSE; a count-appropriate loss does not), but it cannot manufacture an advantage the geometry does not offer.
A positive by-product: the unsupervised latent independently re-derives the hand-built perceptome eigenspace — evidence that the curated coordinate system is real structure in the data — while declining to align with its aging direction, exactly as a pathology-specific framework should.
The honest conclusion for the method: it needs a nonlinearly-encoded target to be worth the machinery. Immune age is the wrong target — and now we know precisely why.
Reproducibility
The full pipeline is pre-registered (three locked amendments), leave-donors-out throughout, fixed seeds, and packaged as a one-click notebook (data download → QC → train ×{real, gene-permuted} → A∧B∧C∧D verdict → learning-order → perceptome cross-check → figures). Every headline number is reported with its matched control and with variance across seeds and folds. Dataset: GSE233321 (healthy human immune system across the lifespan, 167 donors). Code, pre-registration, and the full report — repository in preparation under github.com/mool32.
Part of the perceptome program · methodological negative result · pre-registration discipline per the AI-collaborative research methodology. Theodor Spiro · ORCID 0009-0004-5382-9346 · tspiro@vaika.org